
Le travail d’équipe qui nous a menés jusqu’ici
- Miguel Gomez-Raya Vilanova

- Jun 21, 2024
- 7 min read
Updated: Feb 24
Quelques-unes des grandes questions de la vie sont : d’où vient-on ? Comment en est-on arrivé là ? Pourquoi en est-on arrivé là ? La biologie, à sa manière, essaie de répondre à ces questions. Quelle est l’origine de la vie ? Pourquoi a-t-elle commencé ? Bien qu’il soit difficile de revenir au point de départ, nous pouvons répondre à quelques grandes questions de la vie en étudiant le pourquoi et comment de son évolution. Venez avec moi sur un voyage qui explore un des plus grands événements de la vie sur terre, qui a permis une véritable explosion de la biodiversité : l’origine des eucaryotes.

Tout organisme vivant est composé de cellules. Certains sont multicellulaires, c’est-à-dire composés de plusieurs cellules, comme les animaux et les plantes, et d’autres sont unicellulaires, c’est-à-dire composés d’une seule cellule, comme la bactérie Escherichia coli (1). Cependant, nos cellules individuelles sont très différentes de celles de beaucoup de micro-organismes. D’une part, nos cellules et celles de certains micro-organismes sont très complexes, ayant des compartiments différents, un noyau qui contient le matériel génétique de la cellule, et des organites, comme la mitochondrie, la « centrale énergétique » de la cellule, ou, chez les plantes et les algues, les plastes, qui leur permettent d’obtenir de l’énergie à partir de la lumière. D’autre part, la majorité des micro-organismes sont plus petits et n’ont pas de noyaux. Les biologistes ont utilisé ces différences afin de créer les premières classifications du vivant1 : (i) procaryotes pour les organismes sans noyau (du grec ancien « pró » signifiant « avant » et « káruon » signifiant « noyau ») et (ii) eucaryotes pour ceux avec un noyau (« eu » signifiant « vrai, bien »). Néanmoins, au fil du temps, de nouvelles avancées en biologie moléculaire ont émergé et, avec elles, de nouvelles idées. Dans les années 1970s, Carl R. Woese s’est rendu compte que les procaryotes n’étaient pas tous les mêmes en étudiant l’ARN ribosomique de plusieurs micro-organismes. A première vue, leurs architectures cellulaires étaient, en effet, très similaires, mais l’étude de leurs gènes a démontré que ces organismes étaient très divers (2). Ils pouvaient être classifiés en deux groupes. Depuis, bien que tout organisme vivant soit descendant du dernier ancêtre commun universel (LUCA, pour Last Universal Common Ancestor), on classifie le monde vivant en trois domaines : (i) Bacteria et (ii) Archaea, tous deux des procaryotes, et (iii) Eucarya.
Ces différents types d’organismes ne sont pas apparus en même temps. Les premiers à diverger étaient les Bacteria et Archaea, qui partagent beaucoup de traits, comme leur architecture cellulaire et leurs génomes souvent organisés en un unique chromosome circulaire, avec quelques exceptions. Toutefois, ils ont plusieurs traits divergents, par exemple, la composition de leurs membranes, le mécanisme de réplication de leurs génomes, ou comment leurs informations génétiques sont traitées (3). Cependant, les eucaryotes, plus complexes, apparaissent bien plus tard dans l’histoire de la vie via un processus nommé eucaryogenèse, lorsque le dernier ancêtre commun eucaryote (LECA, pour Last Eukaryotic Common Ancestor) émerge (1). Le processus par lequel LECA a pu acquérir cette complexité cellulaire est un sujet d’actualité chez les biologistes.
Les premier modèles d’eucaryogenèse ont été popularisés par Lynn Margulis (née Alexander), bien que des hypothèses existaient depuis longtemps (1). Elle a fait l’hypothèse que les mitochondries, que l’on sait maintenant présentes dans LECA, et les plastes, trouvés dans les eucaryotes photosynthétiques, aient eu une origine symbiotique. Ceci indiqua que leur origine vienne d’une interaction mutualiste entre deux cellules : un hôte et une cellule qui se changea en organite. Cette dernière, devenue de plus en plus dépendante de la cellule hôte, perdu en complexité, donnant naissance aux organites que l’on trouve aujourd’hui. Margulis a développé cette hypothèse à partir de l’observation que les mitochondries et plastes ont leurs propres ADN et réalisent des fonctions indépendantes de la cellule qui les contient, parmi d’autres caractéristiques. La question était donc : quelles cellules ont donné naissance à ces interactions ? Des analyses de phylogénétique moléculaire de quelques protéines ont pu élucider l’identité des ancêtres probables des mitochondries et plastes1. Leur trouvaille : une alphaprotéobactérie comme cellule hôte et une cyanobactérie comme cellule dépendante. Avec la découverte du domaine Archaea, plusieurs scientifiques se sont demandés si l’hôte aurait été une archée (1),(3).
Les théories actuelles d’eucaryogenèse doivent prendre en compte toutes les données moléculaires et phylogénétiques, acquises grâce à de nombreuses années de recherche. Dernièrement, avec l’avancée du séquençage haut débit, les chercheurs ont réussi à générer une grande quantité de métagénomes, des génomes de micro-organismes parvenant d’échantillons environmentaux (1). La métagénomique renseigne sur une diversité de micro-organismes sans devoir les cultiver en laboratoire, permettant aux scientifiques de passer le cap de l’isolation de ces micro-organismes. Les outils moléculaires, dont la métagénomique, ont contribué aux découvertes de nouvelles lignées et à notre connaissance du monde microbien. Ceci a abouti à la découverte surprenante d’une nouvelle lignée d’Archaea : les archées d'Asgård4. Ceux-ci forment un phylum d’Archaea initialement trouvé dans des sédiments sous-marins profonds. Les premiers génomes de ce groupe sont parvenus de sédiments proches d’une source hydrothermale, nommée le « château de Loki », d’où le nom du premier groupe d’archées d'Asgård (Lokiarchaeota). Toutefois, l’aspect le plus intéressant de cette lignée se trouve dans leur génome. Celui-ci contient l’information génétique pour le codage de protéines crues uniques aux eucaryotes : les protéines de signature eucaryote (4). Ce trait et des analyses phylogénétiques soutiennent l’hypothèse que l’hôte probable de l’eucaryogenèse était, en effet, une archée d'Asgård. Bien qu’il y ait maintenant un consensus sur l’identité de la cellule hôte, grâce à ces nouvelles trouvailles, il existe de nombreuses théories sur les mécanismes et raisons évolutives de l’eucaryogenèse (Figure 1)(1). Dans cet article, nous allons nous concentrer sur deux de ces théories : le modèle « Inside-Out » et la syntrophie.
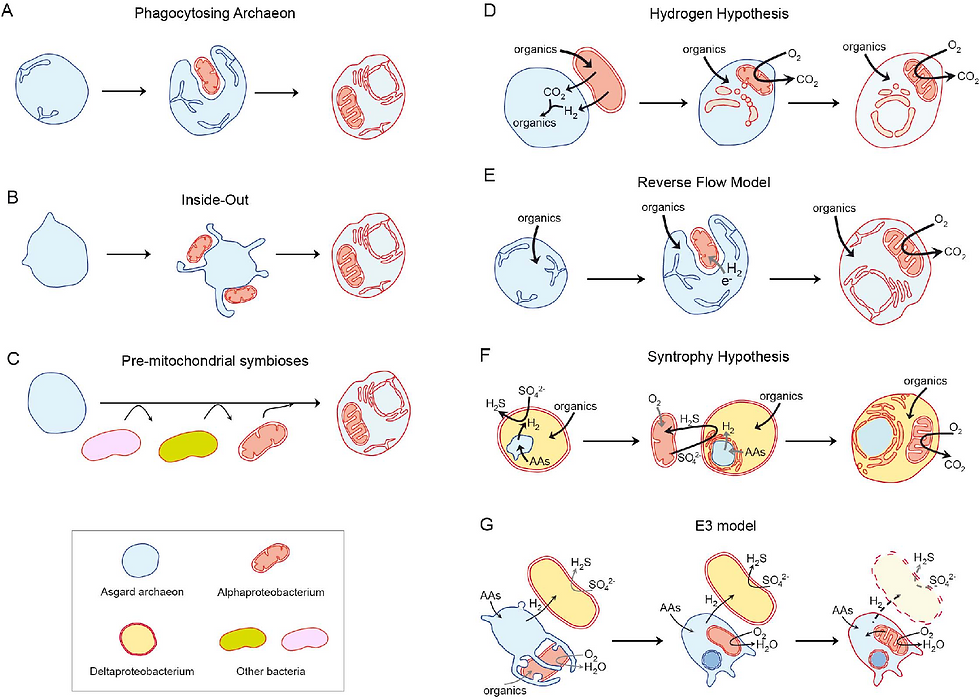
Figure 1
Si l’on créait un eucaryote selon le modèle « Inside-Out » (5), on aurait seulement besoin d’une archée hôte avec la capacité de faire des protrusions dans sa membrane et d’un groupe d’alphaprotéobactéries autour de l’archée. Ces protrusions viendraient ensuite entourer et engouffrer ces bactéries, qui deviendraient par la suite une composante intégrale de la cellule. Ce processus formerait de cette manière les compartiments et membrane internes qu’ont les eucaryotes, ainsi que le noyau. L’isolation et la culture d’une archée d'Asgård, avec imagerie cellulaire montrant la présence de ces protrusions, soutiennent la plausibilité de cette hypothèse (6). Cependant, la composition des membranes des archées est très différente de celle des eucaryotes, elle-même plus proche de celle des bactéries. Donc, comment la membrane a-t-elle changé de composition ? Et, pourquoi ? A ce jour, nous n’avons pas de réponses à ces questions.
D’autre part, si l’on créait un eucaryote selon le modèle de syntrophie (7), nous aurions un scénario très différent. On aurait besoin de trois éléments : une archée d'Asgård, d’une alphaprotéobactérie oxydant le sulfure, et d’une deltaprotéobactérie réduisant le sulfate. Dans ce scénario, l’interaction de ces trois micro-organismes serait maintenue grâce à leurs capacités métaboliques. L’archée produirait de l’hydrogène nécessaire à la réduction du sulfate par la deltaprotéobactérie, entraînant la production de sulfure ensuite utilisé par l’alphaprotéobactérie pour l’oxyder en sulfate, afin de boucler la boucle. Dans ce modèle plus complexe, la deltaprotéobactérie engloutirait d’abord l’archée, qui deviendrait le noyau de la cellule, et puis l’alphaprotéobactérie. L’isolation en laboratoire d’une archée d'Asgård ayant une interaction symbiotique avec une deltaprotéobactérie soutient l’hypothèse de syntrophie (8). De plus, dans ce modèle, la composition de la membrane n’aurait pas à changer puisque la membrane externe serait celle de la deltaprotéobactérie, qui est plus similaire à celle des eucaryotes (9). Néanmoins, si deux événements endosymbiotiques s’étaient déroulés, pourquoi n’avons-t-on pas trouvé des cellules « eucaryotes » sans mitochondrie ou organite dérivé de mitochondrie ? En outre, les chercheurs ont trouvé des gènes dans les génomes eucaryotes d’origine deltaprotéobactérienne, mais aussi d’autres types de bactéries. Donc, y avait-il d’autres bactéries associées à ce processus ?
Pour conclure, le processus complexe d’eucaryogenèse est loin d’être totalement compris. De nouvelles découvertes donnent aux chercheurs une vision plus claire de ce qui aurait pris lieu il y a des milliards d’années. Nous pouvons espérer bientôt avoir des réponses concrètes aux pourquoi et comment de l’eucaryogenèse et enfin éclairer ce mystère qui fascine les biologistes depuis si longtemps.
Réferences
1. López-García, P., and Moreira, D. (2023). The symbiotic origin of the eukaryotic cell. C R Biol. https://doi.org/10.5802/crbiol.118.
2. Woese, C.R., and Fox, G.E. (1977). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.74.11.5088.
3. Lindås, A.-C., and Bernander, R. (2013). The cell cycle of archaea. Nat Rev Microbiol. https://doi.org/10.1038/nrmicro3077.
4. Spang, A., Saw, J.H., Jørgensen, S.L., Zaremba-Niedzwiedzka, K., Martijn, J., Lind, A.E., van Eijk, R., Schleper, C., Guy, L., and Ettema, T.J.G. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature. https://doi.org/10.1038
5. Baum, D.A., and Baum, B. (2014). An inside-out origin for the eukaryotic cell. BMC Biol. https://doi.org/10.1186/s12915-014-0076-2.
6. Rodrigues-Oliveira, T., Wollweber, F., Ponce-Toledo, R.I., Xu, J., Rittmann, S.K.-M.R., Klingl, A., Pilhofer, M., and Schleper, C. (2023). Actin cytoskeleton and complex cell architecture in an Asgard archaeon. Nature. https://doi.org/10.1038/s41586-022-05550-y.
7. López-García, P., and Moreira, D. (2020). The Syntrophy hypothesis for the origin of eukaryotes revisited. Nat Microbiol. https://doi.org/10.1038/s41564-020-0710-4.
8. Imachi, H., Nobu, M.K., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y., Takano, Y., Uematsu, K., Ikuta, T., Ito, M., et al. (2020). Isolation of an archaeon at the prokaryote-eukaryote interface. Nature. https://doi.org/10.1038/s41586-019-1916-6.
9. Krupovic, M., Dolja, V.V., and Koonin, E.V. (2023). The virome of the last eukaryotic common ancestor and eukaryogenesis. Nat Microbiol. https://doi.org/10.1038/s41564-023-01378-y
Cette article a été édité par Kodie Noy. Traduit de l'anglais par Claire Lavergne et Kodie Noy
Rencontrez l'auteur: Miguel Gomez-Raya Vilanova